 |
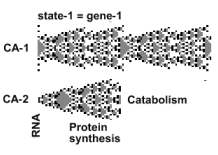
Protein synthesis
 |
Directed turnover
Proteins turn over in a non-random fashion. First the molecule matures, and only then it is catabolized. In a random turnover each state would disappear randomly. .Protein turnover is directed, and obeys the FI-FO rule, First in, is also the first out, .Not only proteins, but all constituents of the organism behave this way, which makes life different from the inorganic world. At any instant, all stages of the evolving protein exist in the body. In the healthy organism, protein formation rate equals its catabolic rate. The organism maintains a steady state, or homeostasis. In order to highlight the fact that all constituents stream, this condition ought to be called homeorhesis (rhesis = stream).
Gene amplification
Each cycle generates one protein molecule. In order to
produce more, CA-1 might cycle somewhat faster. It may also create a new protein assembly line. The
stem process, divides symmetrically,
creating another stem process engaged in protein production,
which is known as gene multiplication.
In order to produce other proteins, CA-1 initiates forms
the CA-2 bud at other states.
To make these arguments more plausible, each state was
flanked by black non-coding sequences (2 = G)
 |
 |
These experiments might imply that CA-1 can generate only transient processes with 46 states. Here is one with 88 states
 |
restoreparams[1,1,1]; restoreparams[2,2,1];
If [stateno[[1]]==35, a[[2]]+=a[[1]]]; go[160]